AGRICULTURAL
COMMUNICATIONS
SERVICES
Providing strategic communications and marketing
for the Arkansas Agricultural Experiment Station
Latest News
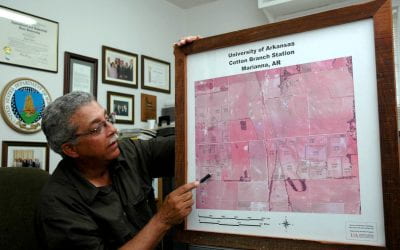
Claude Kennedy, Retired Cotton Station Director, Former ASCS Regional Director, Passes Away at 82
Kennedy, with his radiant smile and outstretched hands, would welcome visitors and instantly make them feel at home. More often than not, a first-time visitor would get a tour and offer of lunch, regardless of what else was on Kennedy’s calendar that day. By Mary...

New Parasite Affecting Canadian Partridges Named for Arkansas Poultry Scientist
Canadian researchers name new Eimeria parasite species after Arkansas scientist By John Lovett – Apr. 16, 2024 NAMESAKE — Billy Hargis, Distinguished Professor of poultry science for the Arkansas Agricultural Experiment...

Food Scientists are Finding Ways to Preserve Food Quality and Ensure Food Safety
Food scientists develop framework to improve food quality and still kill pathogens By John Lovett – Apr. 12, 2024 QUALITY CONTROL — Arshpreet Khattra was the lead author of a study that developed a framework for food...
Claude Kennedy, Retired Cotton Station Director, Former ASCS Regional Director, Passes Away at 82
Kennedy, with his radiant smile and outstretched hands, would welcome visitors and instantly make them feel at home. More often than not, a first-time visitor would get a tour and offer of lunch, regardless of what else was on Kennedy’s calendar that day. By Mary...
New Parasite Affecting Canadian Partridges Named for Arkansas Poultry Scientist
Canadian researchers name new Eimeria parasite species after Arkansas scientist By John Lovett – Apr. 16, 2024 NAMESAKE — Billy Hargis, Distinguished Professor of poultry science for the Arkansas Agricultural Experiment...
Food Scientists are Finding Ways to Preserve Food Quality and Ensure Food Safety
Food scientists develop framework to improve food quality and still kill pathogens By John Lovett – Apr. 12, 2024 QUALITY CONTROL — Arshpreet Khattra was the lead author of a study that developed a framework for food...

USDA-Grant Supported FRST Project Releases New Fertilizer Prediction Tool
Fertilizer recommendation tool for farmers could save money and reduce nutrient loss By John Lovett – Apr. 8, 2024 DECISION AID — Farmers have a new fertilizer decision aid available called FRST (Fertilizer...
Publications
 Research Series 702: Arkansas Soybean Performance Tests 2023
Research Series 702: Arkansas Soybean Performance Tests 2023 Research Series 701: Wayne E. Sabbe Arkansas Soil Fertility Studies 2023
Research Series 701: Wayne E. Sabbe Arkansas Soil Fertility Studies 2023 Research Series 700: Arkansas Cotton Variety Tests 2023
Research Series 700: Arkansas Cotton Variety Tests 2023Research Videos
Meet the Team
We are a team of communications professionals telling the story of the Arkansas Agricultural Experiment Station. Contact us if you have any communications requests. We’d love to hear from you.
MEET THE TEAM >
Resources
Photo Library
Logos
Contact
Have a question? Submit your questions through our contact form.